Halicyclops is a cosmopolitan genus of cyclopoid copepods of the subfamily Halicyclopinae including species and subspecies widely distributed in several kind of surface brackish water bodies, tidal pools, lagoonal and estuarine coasts, anchialine caves, sinkholes, interstitial and sandy beaches habitats, as well as ponds and marshes. The genus is most prolific in the Mediterranean, Caspian, Indian and African regions, with an endemism index about 90%, less known in the North American and Australian areas. However, both the stygobiont and surface copepod fauna of Australia is still under study at present, and first results of our researches (Pesce et al., 1996a, 1996b; Pesce & De Laurentiis, 1996; De Laurentiis et al., in press) are revealing a rich assemblage of species in this area.
The peculiar localizations of some species, such as H. caridophilus Humes, 1947 (Borneo), and the recently described H. tetracanthus Rocha, 1995 (Belize), H. cenoticola Rocha et al., 1998 (Yucatan, Mexico) and H. rochai De Laurentiis et al. (in press) (Australia), lead to link their distribution with the ancient Tethys Sea.
The number of taxa in the genus has been variously recognized, in past times, by numerous authors (Gurney, 1933; Kiefer, 1936; Lindberg, 1957; Wilson, 1958; Herbst, 1977-1987; Defaye & Dussart, 1988; Lotufo & Rocha, 1993; Rocha, 1995; Fiers, 1995). It is so far known from
75 described species and subspecies, most of them inahabiting surface brackish waters, very few species have been reported as exclusive freshwater forms (H. sinensis, H. blachei, H. aberrans, H. lindbergi, H. dussarti), twenty-six are subterranean and they can be found in different grounwater habitats, such as anchialine caves, sinkholes, wells and other various interstitial substrata. Species of Halicyclops can also be found on the body surface or in the tubes of polychaete worms; Halicyclops caridophilus lives in the gill chambers of the mud lobster Thalassina anomala.
Two undetermined species were reported by Plesa (1981) and Suarez et al. (1986) from Cuba and Mexico, respectively. Other questionable registers of European species are not considered herein.
Lists of morphological characteristics to be used in Halicyclops species identification have been given by Kiefer (1936), Lindberg (1949), Wilson (1958) and, more recently, by Rocha (1984) and De Laurentiis et al. (in press). Wilson (1958) and successively Rocha (1995) pointed out that " there is still much to be learned about this genus", and attention should be given, in the future species descriptions, to microcharacters such as the construction and armature of the antenna, antennula and mouthparts, as well as the setation of swimming legs and furcal rami. Taxonomic understanding of the genus has greatly improved during recent years on account of more accurate recognition of the above characters, as well as of formerly overlooked microcharacters, such as morphology and armature of the mouthparts, armature of setae and spines on the distal segment of the endopod of leg 4 and morphology of the hyaline fringe on urosomites, particularly the fourth one.
Particularly, Rocha (1984) called attention to the importance of the ornamentation of apical median setae of the furcal rami, suggesting their consideration in future species descriptions. More recently, the same Author (1993) reported a remarkable dimorphism in the male endopodites of legs 1-2, and the presence of spots ("refractile points") on the dorsal surface of H. maculatus. Lately, De Laurentiis et al. (in press) pointed out the presence of spiniform setae on the paragnaths of H. rochai, up to this time reported only in misophrioid copepods (Huys & Boxshall, 1991), suggesting as well that this character, very likely present also in other cyclopoid copepods, could be somewhat variable and should be more carefully observed and illustrated in species descriptions.
At present, on account of the spine formula of the exopod 3 of legs 1 to 4, the following morphological groups of species can be pointed out:
| |
A - Species with spine formula: 4 4 4 3
B - Species with spine formula: 3 4 4 3
C - Species with spine formula: 3 4 4 2
D - Species with spine formula: 3 4 3 3
E - Species with spine formula: 3 4 3 2
F - Species with spine formula: 2 3 3 3
G - Species with spine formula: 2 3 3 2
| | |
The first group (A) include a single species, H. tenuispina Sewell, 1924, known from brackish waters of Northeastern India (Lake Tchilka). Nevertheless, the placement of this species is rather doubtful since Sewell reported a spine formula "4443", but figured an exopod of leg 4 with 4 external spines (!).
Most species and subspecies fit the second group (B). Within this group a "thermophilus" complex, with the following species, characterized by the presence of a well developed chitinous spiniform process on each side of the genital double-somite, the end of which is curved backward, has been suggested by Herbst (1983) and confirmed by Pesce et al. (1996) and Baribwegure & Dumont (2000) :
H.antiguaensis Herbst, 1983 (West Indies: Bonaire, Antigua)
H.dedeckeri Brownell, 1983 (Cape Town, South Africa)
H.japonicus Ito, 1956 (Japan)
H.latus Shen & Tai, 1964 (China)
H. soqotranus Baribwegure & Dumont, 2000 (Island of Soqotra, Indian Ocean; interstitial)
H.spinifer Kiefer, 1935 (India, Iran, Brazil, Australia)
H.thermophilus Kiefer, 1929 (Java, Madagascar)
H.venezuelaensis Lindberg, 1954 (Venezuela) |
Within the same group a few species possess 8 setae (versus 5 setae in most species) along the lateral margin of the distal endopodal segment of the antenna, but this character has not always been reported in past descriptions. As far as I know, these species are:
H.caneki Fiers, 1995 (Yucatan, Mexico)
H.herbsti Rocha & Iliffe, 1993 (Bermuda)
H.maculatus Rocha & Hakenkamp, 1993 (U.S.A., Brazil) |
The remaining species of this group are characterized by the genital double-somite with or without blunt protuberances directed laterally.
Among these species other useful distinguishing characters could be the length of the distal segment of the antenna, presence/absence of inner seta on endopod segment 3 of leg 4, presence/absence of denticulate hyaline frills on the posterior border of the urosomites, the morphology and the development of the pseudo-operculum, the length of the inner apical furcal setae, the number and length of setae and spines on the exopod of the leg 5, both in males and females, and the construction of leg 6 in both sexes.
H.ambiguus Kiefer, 1967 (Australia)
H.aquae surgentis Bozic, 1964 (Reunion Island)
H.bowmani Rocha & Iliffe, 1993 (Bermuda)
H.caridophilus Humes, 1947 (Borneo)
H.cenoticola Rocha et al., 1998 (Yucatan, Mexico)
H.clarkei Herbst, 1982 (U.S.A.)
H. cf. clarkei Herbst, 1982 (southern U.S.A., Panama, Brazil)
H.coulli Herbst, 1977 (U.S.A.)
H.crassicornis Herbst, 1955 (Brazil)
H.cryptus Monchenko, 1979 (Azov sea)
H.cryptus secundus Monchenko & Polischuk, 1982 (Ukraine; Azov Sea)
H.dalmatinus Petkovski, 1955 (Yugoslavia, Italy)
H.denticulatus Kiefer, 1960 (Madagascar)
H.dussarti Rocha, 1995 (Brazi)
H.electus Lindberg, 1943 (India)
H.exiguus Kiefer, 1934 (Haiti, Costarica, Brazil)
H.gauldi Plesa, 1961 (Ghana)
H.glaber Rocha, 1983 (Brazil)
H.higoensis Ito, 1956 (Japan)
H.hurlberti Rocha, 1991 (U.S.A.)
H.incognitus Herbst, 1962 (Great Britain)
H.konkanensis Lindberg, 1949 (India)
H.korodiensis Onabamiro, 1952 (Africa)
H.laciniatus Herbst, 1987 (Barbados)
H.lindbergi Rocha, 1995 (Brazil)
H.longifurcatus Pesce et al., 1996 (Western Australia)
H.magniceps (Lilljeborg,, 1853) (Europe, Africa, China, North America, Mexico ?)
[ synonyms: H. aequoreus Fisher, 1860; H. christianensis Kiefer, 1935]
H.neglectus Kiefer, 1935 (Europe, Asie, Africa, Madagascar *, central America)
H.oblongus Lindberg, 1951 (Caspian sea) **
H.oraeeburnensis Lindberg, 1957 (Ivory Coast, Brazil)
H.oryzanus Defaye & Dussart, 1988 (Africa)
H.ovatus Rocha, 1984 (Brazil)
H.paradenticulatus Rocha, 1984 (Brazil)
H.pilifer Lindberg, 1949 (India)
H.pilosus Rocha, 1984 (Brazil)
H.pondoensis Wooldridge, 1977 (South Africa, Nigeria ***)
H.propinquus Sars, 1905 (New Zealand) ****
H.reidae Rocha & Hakenkamp, 1993 (U.S.A.)
H.reunionensis Bozic, 1964 (Reunion Island)
H.reunionis Kiefer, 1960 (Reunion Island)
H.robustus Lindberg, 1951 (Caspian sea) **
H.rochai De Laurentiis et al. (in press) (Western Australia)
H.rotundipes rotundipes Kiefer, 1935 (Mediterranean region, Ukraine) *****
H.rotundipes putealis Kiefer, 1938 (south Italy)
H.rotundipes aralensis Borutzky, 1974 (Aral sea)
H. ryukyuensis Ito, 1962 (Japan)
H.septentrionalis (Kiefer, 1935) (Europe, Haiti)
H.similis Kiefer, 1935 (Haiti, French Guiana, Brazil, Panama ?)
H.stocki Herbst, 1962 ( Great Britain)
H.troglodytes Kiefer, 1954 (France, Balearic Islands, Italy, Yugoslavia, Sahara)
H.validus Monchenko, 1974 (Black sea)
H.verae Rocha, 1984 (Brazil)
H.ytororoma Lotufo & Rocha, 1993 (Bermuda, Brazil)
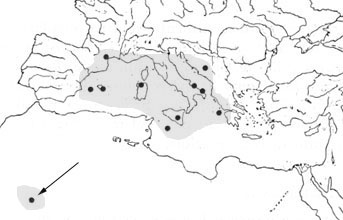
Distribtion of Halicyclops troglodytes Kiefer
* Dussart (1982) pointed out an exceptional individual variation of spine formula in specimens
from Madagascar, viz. 3333, 3343 and 3433 ! ** These species must be allocated to the genus Smirnoviella Monchenko, 1977, forming, together with Smirnoviella unisetosa Monchenko, 1982, the Caspian complex of the genus (Monchenko, in litt.).
*** The register of this species from Nigeria by Boxshall & Braide (1991) is questionable since the Authors figured, as well as reported in their key, the presence of "backward-pointing , curved process" on genital double somite (thermophilus-complex?), on the contrary in the original description of the species the same somite is reported and figured as "genital segment with small protrusions on the anterior lateral margin"; moreover, in the same key the setae on the endopod of leg 4 are reported as normal, but in the original description, and figures, the first endopodal segment of the same leg is armed with a particular, different seta.
**** The systematic status of this taxon is questionable. However, pending a revision of the complex "magniceps-aequoreus" , it should be considered provisionally a valid taxon (Plesa, in litt.).
***** Plesa (1956, 1963), Wilson (1958) and Damian Georgescu (1963) consider H. rotundipes synonym of H. neglectus. Other Authors (Wooldridge, 1977; Monchenko, 1979; Dussart & Defaye, 1985; Rocha, 1995) do not share this opinion.
The group "C" include three species, viz. H. fosteri Wilson, 1958, known from Lousiane (U.S.A.), and H. setifer Lindberg, 1950 and H. sarsi Akatova, 1935, both from Caspian sea. However, the last species, after a more accurate examination, revealed to be a senior synonym of H. setifer (Monchenko, in litt.).
At the group "D" belong five taxa: H. brevispinosus meridionalis Herbst, 1953, from Romania, Black sea, Azov sea; H. laminifer Herbst, 1982, from U.S.A.; H. tetracanthus Rocha, 1955, from Belize; H.sinensis (Kiefer,1928), from China and Japan; H. exiguus sensu Defaye & Dussart, 1988 (French Guyana) [ In my opinion, and according to Rocha (in litt.), this last taxon must represent a new species].
A single species, H. blachei Lindberg, 1952 (Cambogia) fits the group "E".
The group "F" is represented by four species: H. brevispinosus Herbst, 1952 (Germany), H. tageae Lotufo & Rocha, 1993 ( Brazil), H. canui Lindberg, 1941 (India) and H. pusillus Kiefer, 1954 (Madagascar).
The last group (G) includes a single species, viz. H. aberrans Rocha, 1983, known for Brazil and French Guiana. Wilson (1958) reported the formula "2332" as an exception in H. fosteri.
As regard the armature of the exopod of leg 5, and independently of spine formula of exopod 3 of legs 1 to 4, a "caridophilus" group has been recently suggested for those species having a male leg 5 exopod armed with 3 spines and 3 setae (De Laurentiis et al., in press); to this group. at present, belong the following species: H. caridophilus, H. cenoticola, H. rochai, H. tetracanthus, but for many species males are unknown or undescribed. Other species (H. aberrans, H. crassicornis, H. denticulatus, H. exiguus sensu Kiefer, H. gauldi, H. maculatus and H. tageae) are characterized by the same male exopod not sexually dimorphic, armed with four appendages.

DE LAURENTIIIS P., G.L.PESCE & W.F. HUMPHREYS. 1999. Copepods from ground waters of Western Australia. IV. Cyclopids from basin and craton aquifer (Crustacea: Copepoda: Cyclopidae). Rec. West. Austr. Mus., 19: 243-257.
FIERS F. 1995. Halicyclops caneki n. sp. (Copepoda, Cyclopoida) from Celestun lagoon (Yucatan, Mexico). Belg. J. Zool., 125 (2): 301-303.
LOTUFO G. R. & C. E. F. ROCHA. 1993. Intertidal interstitial Halicyclops from the Brazilian coast (Copepoda: Cyclopoida). Hydrobiologia, 264: 175-184.
MIRABDULLAYEV, I.M. & I.A. GETZ. 1996. Halicyclops spinifer (Kiefer, 1935) (Crustacea, Copepoda) - a tropical species, new to the fauna of Central Asia. Doklady Akademii Nauk Respubliki Uzbekistan, 4:43-44.
PESCE G. L., P. DE LAURENTIIS & W.F. HUMPHREYS. 1996. Copepods from ground waters of Western Australia. II. The genus Halicyclops (Crustacea: Copepoda: Cyclopidae). Rec. West. Austr. Mus., 18: 77-85.
ROCHA C.E.F. 1991. A new species of Halicyclops (Copepoda, Cyclopidae) from California, and a revision of some Halicyclops material in the collections of the US Museum of Natural History. Hydrobiologia, 226: 29-37
ROCHA C.E.F. 1995. Copepods of the genus Halicyclops (Cyclopidae) from Belize. Hydrobiologia, 308: 1-11
ROCHA C.E.F. 1995. Two new species of Halicyclops (Copepoda, Cyclopoida) from the Amazon Basin, Brazil. Hydrobiologia, 315: 167-175.
ROCHA C. E. F. & M. ILIFFE. 1993. New cyclopoids (Copepoda) from anchialine caves in Bermuda. Sarsia, 78:43-56.

HERBST H.V. 1982. Drei neue Cyclopoida Gnathostoma (Crustacea:Copepoda) aus dem nordamerikanischen Küstenbereich. Gewässer und Abwässer, 68/69: 107-124.
LINDBERG K. 1957. Cyclopides (Crustacés copépodes) de la Cote d'Ivoire.. Bull. I.F.A.N., 19, ser.A (1):134-179.
REID J. 1985. Chave de identificaçăo e lista de referęncias
bibliográficas para as especies continentais sul-americanas de vida
livre da ordem Cyclopoida (Crustacea, Copepoda). Bolm Zool, Univ. S.
Paulo 9: 17-143.
ROCHA C.E.F. & C. C. HAKENKAMP. 1993. New species of Halicyclops (Copepoda Cyclopidae) from the United States of America. Hydrobiologia, 259: 145-156.
ROCHA C.E.F. , T.M. ILIFFE, J.W. REID & E. SUAREZ-MORALES. 1998. A new species of Halicyclops (Copepoda:Cyclopoida:Cyclopidae) from cenotes of the Yucatan Peninsula, Mexico, with an identification key for the species of the genus from the Caribbean region and adjacent areas. Sarsia, 83:387-399.
WILSON M. S. 1958. The copepod genus Halicyclops in North America, with description of a new species from Lake Pontchartrain, Louisiana, and the Texas coast. Tulane Studies in Zoology, 6 (4): 176-189.
WOOLDRIDGE T. 1977. A new species of Halicyclops (Copepoda, Cyclopoida) from estuaries in Transkei, Southern Africa. Ann. S. Afr. Mus., 73 (10): 361-371.
DE LAURENTIIS PAOLA
Dipartimento di Scienze Ambientali - Universitŕ di L'Aquila
Via Vetoio - 67100 - L'Aquila (Italy)
FIERS FRANK
Royal Belgian Institute of Natural Sciences
Vautierstraat 29, B-1000 Brussels - Belgium
E-mail: frankfiers@kbinirsnb.be
HERBST H.V.
Distelweg 1
4150 Krefeld 29 - Germany
ILIFFE THOMAS M.
Department of Marine Biology - Texas A&M University at Galveston
Galveston, TX 77553-1675
Phone: 409 740-4454 Fax: 409 740-5001
e-mail: iliffet@tamug.tamu.edu
http://www.marinebiology.edu/iliffe.html
MONCHENKO V.
Schmalhausen Institute of Zoology - Ukrainian Nat. Acad. of Sciences
B.Khmelnits'kogo 15, Kiev-30, UKRAINE
Tel: (380-44) 225-10-70;: Fax: (380-44) 224-15-69
e-mail: root@iz.freenet.kiev.ua
PESCE GIUSEPPE L.
Dipartimento di Scienze Ambientali - Universitŕ di L'Aquila
Via Vetoio - 67100 - L'Aquila (Italy)
tel. 0039-862-433232; Fax: 0039-862-433205
e-mail: giuseppe.pesce@aquila.infn.it
http://www.univaq.it/~sc_amb/pesce.html
http://www.glpesce.net
PLESA CORNELIU
Str. Gh. Dima 28/27
3400 Cluj-Napoca 6 - Roumanie
Fax: 0040-64-195954;
e-mail:plesa@mailexcite.com
REID JANET W.
Department of Invertebrate Zoology
MRC-163 National Museum of Natural History - Smithsonian Institution
Washington DC 20560- USA
tel. (202) 357-4674 fax (202) 357-3043
e-mail: 74024.1511@compuserve.com
ROCHA CARLOS E.
Departamento de Zoologia IBUSP
Caixa Postal 11461 - 05422-970 Sao Paulo, Brazil
Fax: 0055-11-8187416; e-mail: cefrocha@usp.br
SUAREZ-MORALES EDUARDO
ECOSUR, El Colegio de Frontera Sur
Unidad Chetumal, Apdo Postal 424
Chetumal, Q. Roo - 77000 Mexico
 LINK TO THE COPEPOD WORLD AT SMITHSONIAN INSTITUTION LINK TO THE COPEPOD WORLD AT SMITHSONIAN INSTITUTION
Sincere thanks are due to V.Monchenko, H.V.Fiers, C.Plesa and
C.E.Rocha for useful suggestions and/or improvementsPlease, for comments and/or suggestions e-mail to: pesce@univaq.it |
[HOME] [BACK] |
|