|

Il subphylum degli Uniramia è suddiviso in due classi: INSECTA e MYRIAPODA. Gli unirami moderni sono prevalentemente artropodi terrestri,gli unirami acquatici si riadattati successivamente a tali habitat. La classe degli insetti è la più variegata per forme ed adattamento ad ogni possibile habitat terrestre,d'acqua dolce e marini. Gli insetti rappresentano un importante componente della dieta di molti animali e sono fondamentali per la biosfera in quanto molti di essi agiscono come decompositori;inoltre,svolgono un ruolo predominante nell'impollinazione delle piante a fiore;alcuni insetti sono anche di interesse economico poichè producono miele,cera,seta.
L'anatomia degli Unirami è il risultato di molti adattamenti alla vita in ambienti terrestri, come ad esempio appendici di sostegno e locomozione più forti e efficienti, adattamenti per sopportare stress osmotici e strutture aeree per gli scambi gassosi. Gli insetti, come gli aracnidi, hanno evoluto strutture di protezione contro la perdita di acqua. Il corpo degli Unirami è composto da tre o due tagmi rispettivamente negli Insetti e nei Miriapodi; capo e tronco plurisegmentato nei Miriapodi; capo, tronco e addome plurisegmentato negli Insetti. Le appendici sono multiarticolate ed uniramose, le appendici del capo sono: antenne, mandibole, primo paio di mascelle, secondo paio di mascelle, queste ultime sono spesso fuse a formare un labium. I tre segmenti del torace degli insetti vengono chiamati protorace, mesotorace, metatorace. Ogni segmento è suddiviso in quattro regioni composte da sleriti, detti: noto dorsale, sterno ventrale a sua volta costituito da uno sternite e da due pleuriti laterali. Gli scleriti vengono nominati in base al segmento in cui si trovano, ad esempio gli scleriti del protorace sono detti pronoto, prosternite e propleuriti. I tre segmenti toracici presentano ciascuno un paio di zampe ambulacrali articolate sui pleuriti. Le ali, quando sono presenti si articolano sul noto o sui pleuriti del mesotorace e del metatorace. L'addome è suddiviso in undici segmenti, alcuni segmenti possono fondersi; i segmenti addominali comprendono due scleriti, il tergite e lo sternite.
L'addome non presenta appendici, anche se esistono abbozzi nello stadio larvale, in molti insetti vi sono due appendici sull'ultimo segmento detti cerci, questi hanno funzione sensoriale. Gli insetti presentano uno strato ceroso nell'epicuticola, questo strato ha permesso loro di sopravvivere anche negli ambienti più secchi. I miriapodi hanno strutture molto meno specializzate rispetto agli insetti, la metameria è molto più evidente, soprattutto nelle strutture interne, inoltre non hanno un torace differenziato. Le appendici del capo e le zampe sono simili a quelle degli insetti.
Come negli altri artropodi il capo è formato da un acron presegmentale più un numero di metameri fusi. Nell'acron si trovano gli occhi ed il protocerebro. Il primo metamero cefalico è detto segmento antennale, porta le antenne ed ospita il deutocerebro. 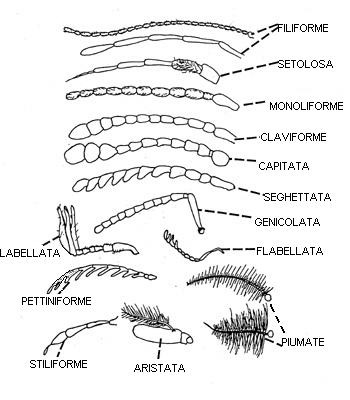
Il secondo metamero è detto segmento premandibolare che non porta appendici, contiene il tritocerebro. Il terzo metamero è detto segmento mandibolare perché porta le mandibole. Il quarto ed il quinto sono detti rispettivamente primo segmento mascellare e secondo segmento mascellare, le seconde mascelle sono fuse a formare un labium nei sinfili e negli insetti. Gli unirami presentano due ulteriori strutture del capo. Sul lato frontale del capo, anteriormente alla regione boccale si trova una piastra detta labrum o labbro superiore, questa va a coprire le mandibole. Negli insetti il labrum è attaccato ad uno sclerite detto fronte dotato di un ocello. Posteriormente al campo boccale, dietro le prime mascelle c'è una struttura lembiforme detta ipofaringe, attraverso cui sfociano le ghiandole salivari. L'orientamento del capo e delle parti boccali negli insetti è variabile rispetto al resto del corpo. Nella condizione ipognata le parti boccali occupano la zona posteriore del capo in continuità con le zampe, è la condizione più primitiva, in genere è presente nei gruppi erbivori e succhiatori di sangue.

Nella condizione prognata, invece, la bocca e le parti boccali sono spostate in avanti, questa condizione è prevalente nelle specie carnivore. Negli insetti degli ordini Hemiptera ed Homoptera le parti boccali sono opistorinchi, formano una proboscide allungata rivolta indietro tra le zampe posteriori.
Gli unirami hanno un esoscheletro sclerotizzato adatto a sostenere il corpo e gli arti sulla terraferma. 
Negli unirami i movimenti antero-posteriori degli arti si hanno tra la coxa ed il corpo propriamente detto. 
La velocità di deambulazione è variabile nei diversi ordini; nei millepiedi in genere le zampe sono corte perché adattate ad un'andatura lenta e potente; negli unirami in grado di correre, come molti insetti, centopiedi e sinfili, gli arti sono lunghi e perciò sono capaci di coprire una maggiore angolazione; nei pauropodi si va da specie dette aratori lenti ad altre specie di corridori. Altri adattamenti degli arti e dei meccanismi di locomozione si hanno soprattutto in risposta ai comportamenti alimentari. Molti centopiedi sono carnivori e perciò tendono a correre velocemente: molti millepiedi, invece, sono prevalentemente erbivori o spazzini e si affossano nel terreno e per tale ragione sono lenti e potenti. I millepiedi ed i pauropodi quando si affossano ripiegano il capo ventralmente in modo che il collo venga spinto in avanti come una pala. Esiste un gruppo di chilopodi, e gli organismi che gli appartengono sono detti geofilomorfi, questi si affossano grazie alla dilatazione del corpo e della muscolatura del tronco, con movimenti peristaltici simili a quelli dei lombrichi; questo può avvenire nonostante l'esoscheletro rigido per la presenza di estese regioni di cuticola flessibile. La lunghezza degli arti comporta che il campo di movimento di un arto possa sovrapporsi a quello di un arto adiacente. Questa interferenza, negativa per l'equilibrio, viene impedita da una diversa lunghezza degli arti. Gli insetti si muovono con una sequenza tripode alternata, l'equilibrio viene mantenuto poiché tre zampe rimangono sempre a contatto del terreno.
Il volo è una tra le più importanti conquiste degli insetti, che sono gli unici invertebrati propriamente volatori. Vi sono numerosi insetti atteri, questi però appartengono a gruppi che hanno perso le ali (pulci, pidocchi) oppure a gruppi primitivi (colemboli, tisanuri). Le ali sono poste sul secondo e sul terzo segmento toracico. Negli ordini dictera e coleoptera c'è un solo paio di ali, nei coleotteri l'altro paio forma coperture dorsali protettive (elitre), nei ditteri formano delle strutture di equilibrio, utili per il volo (bilancieri). Attualmente non si sa con certezza quale sia stato il percorso evolutivo che ha portato alla nascita di insetti alati, questa incertezza è dovuta alla difficoltà di rintracciare le fasi intermedie.
Gli insetti alati più primitivi (odonati, ortotteri…) hanno ali membranose a ventaglio sostenute da molte vene che formano una fitta rete, in molti fossili sono state rinvenute delle strutture simili. Col progredire degli studi hanno preso piede due teorie; quella di più successo è la teoria paranotale secondo cui le ali sarebbero derivate da pliche laterali dei noti toracici con funzioni di stabilizzatori, sono state rinvenute pliche del genere in alcuni insetti fossili. L'altra teoria è quella branchiale, ipotizza che gli insetti pterigoti siano derivati da insetti acquatici e quindi che i precursori delle ali fossero branche toraciche con funzione di stabilizzatori; queste si sarebbero successivamente specializzate come veri e propri arti natatori. Negli insetti moderni le ali sono pieghe dell'integumento le cui superfici, superiore ed inferiore, sono formate da sottili membrane cuticolari. Le vene che attraversano le ali contengono emolinfa circolante. 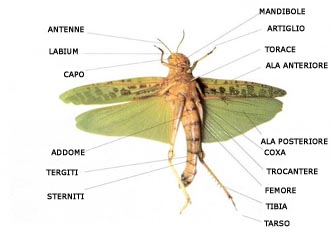
Gli insetti volano battendo le ali creando correnti d'aria da cui traggono portanza, il meccanismo di volo è molto complesso e lontano da quello di uccelli e pipistrelli.
L'ala è articolata col bordo del noto, la sua estremità prossimale posa su di un processo pleurale che funge da fulcro, l'articolazione è costituita dalla proteina elastica: resilina. I movimenti di orientamento delle ali sono molto vari e consentono anche manovre complesse, tali manovre sono rese possibili anche grazie alla flessibilità dell'ala e all'azione di diversi complessi muscolari. I muscoli diretti del volo sollevano e abbassano le ali e ne inclinano il piano a varie angolazioni. La forza della battuta proviene da due gruppi di muscoli indiretti del volo. 1. muscoli dorsali longitudinali, si trovano tra gli apodemi all'estremità posteriore ed anteriore del segmento alato, quando si contraggono provocano un accorciamento del segmento e di conseguenza l'abbassamento delle ali;
2. muscoli dorso ventrali, vanno dal noto allo sterno nei segmenti alati e sono antagonisti dei precedenti, contraendosi provocano l'accorciamento dorso-ventrale del segmento e quindi determinano il sollevamento delle ali. Si comprende così che sono le deformazioni dell'esoscheletro e le sue proprietà elastico-meccaniche a generare il battito di ali.
Il rimbalzo elastico che si ha dalla deformazione, contribuisce allo stimolo alternato dei muscoli antagonisti.
Negli uniramia si hanno svariati comportamenti alimentari, vi sono organismi carnivori, erbivori, spazzini e vari tipi di simbiosi. I centopiedi sono prevalentemente predatori attivi di invertebrati ed altri artropodi, i loro massillipedi sono strutture ingrandite a formare artigli o prensilli che servono per ferire e per iniettare il veleno.
I centopiedi geofiliformi possiedono delle ghiandole repellenti con funzione difensiva poste sugli sterniti. I millepiedi sono erbivori che si nutrono di materiale vegetale morto ed in decomposizione e perciò sono importanti in senso ecologico; alcune specie tropicali sono specializzate per nutrirsi di succhi vegetali.
Esistono alcuni gruppi di millepiedi predatori come i centopiedi che sono anche dotati di ghiandole repellenti. I pauropodi sono generalmente spazzini, strisciano nel terreno nutrendosi di funghi, materiale vegetale e animale in decomposizione. I sinfili sono soprattutto erbivori, ma alcuni gruppi sono anche carnivori o spazzini. Gli insetti, invece, mostrano una vastissima varietà di strategie nutrizionali, in base ad esse gli insetti vengono classificati come: pungenti-masticatori, succhiatori, lambenti. 1. I pungenti-masticatori (es. le cavallette) hanno le parti boccali meno specializzate, le mascelle ed il labium presentano palpi sviluppati simili a zampe, questi servono a tenere fermo il cibo mentre viene masticato e lacerato dalle mandibole. Queste ultime sono prive di palpi ed hanno piccoli denti aguzzi, mentre le appendici scorrono una contro l'altra. Gli insetti con tale apparato boccale possono essere carnivori, erbivori o spazzini, sono però prevalentemente consumatori di piante verdi. 2. I succhiatori sono specializzati nel consumo di linfa vegetale, nettare, sangue o liquidi cellulari, tale strategia alimentare si è evoluta ripetutamente in vari ordini. In alcuni succhiatori, come ad esempio le zanzare, la nutrizione avviene con la perforazione del tessuto animale vivo e poi con l'aspirazione del sangue, sono pertanto perforanti-succhiatori. I lepidotteri (farfalle) si nutrono del nettare dei fiori, sono esclusivamente succhiatori, le parti boccali sono allungate a formare una proboscide aghiforme. Altri insetti, come i miridi e gli emitteri sono perforanti-succhiatori ed hanno un becco costituito dal labium a forma di mangiatoia, da due mandibole e da due mascelle appuntite. Le pulci hanno tre stiletti, il labrum e le due mandibole. Nelle api le prime mascelle ed il labium sono modificati in modo da formare un tubo per succhiare il nettare: le mandibole sono conservate e vengono utilizzate per manipolare la cera durante la costruzione del nido. I succhiatori possono avere altre strutture associate alle parti boccali, queste servono per assorbire altri liquidi; vi può essere una pompa alimentare evolutasi con l'allungamento della cavità orale o cibaria; nei lepidotteri, ditteri ed imenotteri la pompa cibariale è collegata con una pompa faringea, associate a questa vi sono varie ghiandole salivari utilizzate per ammorbidire i tessuti della preda. 3. I lambenti, ovvero la maggior parte dei ditteri (es. mosca domestica) hanno un labium esteso a formare una struttura porosa detta labello, i liquidi vengono trasportati attraverso capillari che vanno dal labello alla bocca.
Nelle forme lambenti sono assenti le mandibole, tali appendici sono conservate soltanto nei perforatori-lambenti (es. mosca cavallina). 4. Molti insetti sono simbionti e parassiti, le loro strategie simbiotiche sono varie. Vi sono tre ordini (Mallophaga, Anaplura, Siphonoptera) composti quasi totalmente da parassiti atteri che trascorrono l'intero ciclo vitale su di un ospite. I Mallofagi parassitano uccelli e mammiferi; hanno un capo largo con parti boccali pungenti atti a maciullare le cellule epiteliali dell'ospite. Gli anapluri sono anch'essi pidocchi parassiti di mammiferi (uomo compreso): hanno un capo stretto con parti boccali perforanti-succhiatrici. Le uova dei pidocchi o lendine vengono attaccate ai peli ed alle piume dell'ospite, da queste emergono i piccoli che passano direttamente allo stadio adulto.
Le pulci (siphonoptera) sono parassiti di mammiferi; a differenza dei pidocchi sono olometabole, subiscono la metamorfosi e quindi passano attraverso gli stadi di uovo, larva, pupa, adulto. Le uova non vengono depositate sull'ospite, ma nell'ambiente, inoltre, le larve si nutrono di detriti organici. Altri ordini comprendono insetti a vita libera, di questi però, alcune specie sono parassite o micropredatrici, altre hanno stadio larvale parassita e stadio adulto a vita libera.
Molti di questi ordini si trovano a metà strada tra il parassitismo obbligato e la predazione, per tale motivo vengono classificati come insetti parassitoidi. Le cimici del letto si nutrono del sangue dell'ospite, ma non vivono su di esso; hanno parti boccali simili agli anopluri. Varie specie di insetti sono vettori di malattie (es. le zanzare che trasmettono la malaria, altri insetti sono vettori della tripanosomiasi o morbo di Chagas). Esistono anche simbiosi mutualistiche tra formiche (ordine Hymenoptera) e piante; le piante si sono adattate a crescere con le formiche per poter sopravvivere in terreni poveri di sostanze nutritive.
Le formiche forniscono alle piante nutrimento, protezione e pulizia in cambio di cibo e rifugio.
Un esempio di tale simbiosi è la relazione tra Pseudomyrmex ferruginea e acacia cornigera. Altri imenotteri sono responsabili di strategie parassite, alcune vespe depositano le loro uova all'interno dell'esoscheletro di altri artropodi,in tal modo le larve potranno crescere nutrendosi dei tessuti dell'ospite fino al raggiungimento dello stadio adulto . In genere i coleotteri sono vittime di questa tattica, ma la specie Peps limbata è in grado di deporre le proprie uova all'interno dell'esoscheletro delle tarantole dopo avervi iniettato un veleno paralizzante.
|


