
Nel 1882 Walther Flemming individua i filamenti che si separano in una cellula al momento della sua divisione e duplicazione, distribuendosi successivamente nelle due cellule-figlie: č la scoperta dei cromosomi, che verranno cosě denominati nel 1888 da Heinrich Wilhelm Waldeyer. Successivamente, nel 1902 il genetista americano Walter Stanford Sutton (1877-1916) ipotizza che i cromosomi siano i "contenitori" dei fattori ereditari che si trasmettono alla discendenza e che gli stessi sarebbero formati da "collane" di elementi di base dei caratteri ereditari, che il botanico danese Wilhelm Ludwig Johansen (1857-1927) per la prima volta chiama geni. Morgan nel 1910 compie ricerche sulla Drosophila melanogaster, il moscerino della frutta che č ancora oggi il materiale elettivo delle ricerche di genetica grazie alla semplicitŕ del suo corredo genetico e alla notvole,velocitŕ con cui si riproduce. Morgan stabilisce che esistono particolari cromosomi che trasmettono i caratteri sessuali (cromosoma X per le femmine; Y per i maschi) e ottiene la prima mappa cromosomica della Drosophila. Nel 1915, Morgan e i suoi collaboratori pubblicarono un testo in cui esponevano chiaramente l’ipotesi che i fattori mendeliani sono i geni, particelle materiali localizzate sui cromosomi, ed a questo autore ed ai suoi seguaci si deve la definizione dei principi fondamentali della moderna teoria cromosomica dell’ereditarietŕ. A partire dal 1927 si inaugura ufficialmente il grande filone di ricerca sull'ereditarietŕ che, combinando alcune scoperte precedenti sul materiale genetico e la teoria della trasmissione dei caratteri ereditari porterŕ, negli anni successivi, alla scoperta della struttura del Dna e, in tempi piů recenti, alla definitiva affermazione della genetica e della biologia molecolare.
I cromosomi, composti da proteine e DNA, sono corpi densi che si trovano all'interno del nucleo delle cellule. L'informazione genetica č contenuta nel DNA dei cromosomi sotto forma di sequenze lineari di basi (A,T,C.G). Il DNA di un singolo cromosoma č una lunga molecola, spiralata e altamente condensata. Il numero complessivo di basi in tutti cromosomi di una cellula umana č circa 6 bilioni (!) e ogni singolo cromosoma contiene da da 50 a 250 milioni di basi. Ogni cromosoma puň contenere diverse migliaia di geni e poche o milioni di basi. 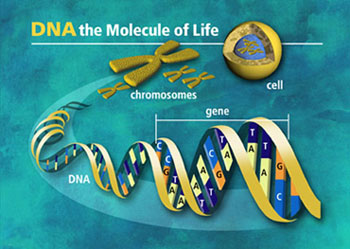
Ogni cromosoma č costituito da una sequenza di geni, ognuno dei quali determina un carattere diverso. Nel 1944 si scoprě che il DNA č il materiale di cui sono costituiti i geni, e che tutta l’informazione necessaria alla continuazione della vita, di generazione in generazione, č inclusa in questa molecola. Un gene č, quindi, un tratto di DNA, responsabile della determinazione di un dato carattere e puň presentarsi in piů forme diverse: le diverse forme dello stesso gene vengono definite dette "alleli". Nel fenomeno della pleiotropia un singolo gene puň presentare diverse espressioni fenotipiche.  Per ciascun gene, responsabile di un particolare carattere, nel nucleo di tutte le cellule somatiche di ogni individuo sono presenti due copie, una di provenienza materna ed una paterna; ciascuna copia si trova nella stessa posizione (locus genico), su ciascuno dei due cromosomi omologhi. Le due copie di ogni allele si separano durante la formazione dei gameti perché i due cromosomi di una coppia di omologhi si separano durante l’anafase della meiosi. Successivamente l’ unione dei gameti, al momento della fecondazione riunisce due corredi genici. Quando i due allleli di un gene sono identici, l’individuo č definito omozigote per quel particolare gene. Molto spesso, tuttavia, queste due copie possono essere diverse: ad esempio, quando un genitore ha gli occhi azzurri e l’altro ha gli occhi marroni, nella discendenza possono essere presenti due copie diverse del gene, una per ciascun colore, una trasmessa dal padre e l’altra dalla madre: in questo caso l’individuo č eterozigote per quel gene. Nelle cellule somatiche per ogni gene sono, quindi, presenti due alleli, di cui generalmente solo uno dei due č manifesto e maschera l’altro; il primo viene detto "allele dominante", il secondo "allele recessivo". Il carattere recessivo puň manifestarsi, nelle generazioni successive, negli individui omozigoti per il gene considerato. La combinazione dei geni e degli alleli che un individuo possiede ed č in grado di trasmettere alle successive generazioni costituisce il genotipo individual; con il termine fenotipo si indica, invece, l’insieme delle caratteristiche morfo-fisiologiche esteriori di un individuo. A volte, ma non sempre, il fenotipo riflette il genotipo: infatti se un individuo possiede due alleli recessivi o due alleli dominanti per un dato carattere, il fenotipo corrisponde al genotipo, ma se un individuo possiede un allele dominante e uno recessivo, il fenotipo č quello del carattere dominante, e di conseguenza rimane nascosto l’allele recessivo. Per ciascun carattere ereditario sono possibili tre genotipi diversi, corrispondenti a due fenotipi:
L’allele dominante si manifesta a livello fenotipico sia nell’individuo omozigote dominante, sia nell’eterozigote; quello recessivo si manifesta invece solo nell’omozigote recessivo.
I geni situati sullo stesso cromosoma vengono definiti "associati", cioč sono allineati in un' unica fila e generalmente vengono trasmessi "a blocchi" (Legge della disposizione lineare dei geni). Per esempio, nella Drosophila una stessa coppia di cromosomi che porta i geni relativi alla forma delle ali porta anche i geni relativi al colore degli occhi. Il suddetto allineamento genico puň, tuttavia, venire alterato da un particolare processo che si verifica durante la meiosi: il "CROSSING OVER". 
Tale fenomeno, scoperto dallo zoologo americano T. H. Morgan, consiste nel fatto che alla meiosi i cromosomi appaiati possano in alcuni casi "attorcigliarsi" l'uno con l'altro (crossing over) e rompersi successivamente nel punto di incrocio; le parti staccate potrebbero, quindi, riunirsi in modo "sbagliato". Questo fenomeno, quando si verifica, comporta la produzione di gameti prevedibili ed, in misura minore, di gameti non prevedibili, con diverso assortimento allelico. 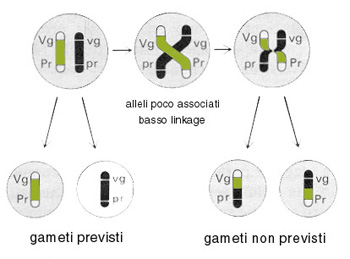
La frequenza degli "scambi", cioč di "crossing-over", č direttamente proporzionale alla distanza tra due geni, cioč del loro "linkage". Se, cioč, due geni sono molto vicini l'uno all'altro, le probabilitŕ che lo scambio si realizzi proprio tra tali punti ravvicinati risulterŕ molto scarsa; al contrario, se i due geni sono piuttosto distanti la probabilitŕ di scambio tra di essi risulta piů elevata. 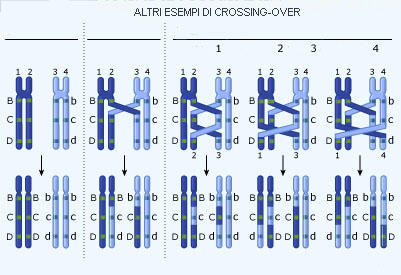
All Rights Reserved. Text and images on this website may not be redistributed or put |


