Per sviluppo embrionale si intende il processo che consente allo zigote di accrescersi, differenziarsi ed acquisire le caratteristiche della specie cui appartiene. Perchè questo processo possa aver luogo devono essere soddisfatte due condizioni fondamentali: un ambiente favorevole che impedisca la disidratazione, e la disponibilità di riserve energetiche. Le prime fasi dello sviluppo sono influenzate dal tipo di uovo di partenza. A questo riguardo le uova si classificano in base alla quantità di materiale di riserva (tuorlo) in esse contenuto:
uova a-lecitiche, tipiche dei Mammiferi Euteri, sono prive di tuorlo e vanno incontro a divisione totale.
uova oligolecitiche che contengono poco tuorlo; le più comuni sono le uova dei mammiferi, degli anfiossi e dei ricci di mare. L'embrione si trova presto nelle condizioni di non dipendere dall'uovo per sopravvivere.
uova mesolecitiche con media quantità di tuorlo. Le più comuni sono le uovadei Ciclostomi Petromizonti, degli Osteitti Attinopterigi Condrostei ed Olostei e degli anfibi. Il sostentamento dell'embrione viene garantito solo per le prime fasi dello sviluppo.
uova telolecitiche con grande quantità di tuorlo. Le più comuni sono le uova dei Ciclostomi Mixinoidei, dei Condroitti, degli Osteitti Attinopterigi Teleostei e degli Osteitti Sarcopterigi, dei Rettili, degli Uccelli e dei Mammiferi Prototeri. La notevole quantità di tuorlo garantisce il sostentamento per tutto il periodo dello sviluppo. Il tuorlo (deutoplasma) dell'uovo in genere non è distribuito in modo uniforme, ma secondo un gradiente di concentrazione: è più abbondante verso il polo vegetativo dell'uovo, mentre il citoplasma si trova concentrato al polo opposto (polo animale). L'asse immaginario che unisce i due poli è detto asse polare. Il deutoplasma delle uova meso e telolecitiche si dispone secondo un gradiente di concentrazione lungo l'asse che unisce i due poli. La polarità dell'uovo, già manifesta nel corso dell'ovogenesi, si esprimerà poi nel piano generale di organizzazione dell' embrione.  Lo sviluppo embrionale si suddivide in tre fasi principali: la segmentazione, la gastrulazione e l'organogenesi. La segmentazione inizia subito dopo la fecondazione: l' uovo fecondato (zigote) comincia a dividersi. All' inizio va incontro ad una serie di numerose divisioni mitotiche che portano alla formazione di una morula e, successivamente, di una blastula: questo processo è sotto il controllo dell RNA fornito dall'uovo. Si conoscono due tipi principali di segmentazione: segmentazione totale e segmentazione parziale. La segmentazione totale, o oloblastica, coinvolge tutta la cellula dello zigote; la segmentazione parziale, o meroblastica, coinvolge solo una parte della cellula zigotica; questo avviene, generalmente, poiché c’è una gran presenza di vitello o deutoplasma all’interno della cellula nel momento dell’accrescimento cellulare (auxocitosi).
In questi casi intervengono processi di segmentazione parziale che si suddividono in discoidali e superficiali. La segmentazione parziale discoidale si osserva solamente nei Cefalopodi e in parte dei Cordati e si segmenta una piccola zona, a forma lenticolare, che contiene buona parte del plasma formativo. La zona ricca di plasma è situata, solitamente, all’apice dell’uovo. La segmentazione inizia con la cariodieresi, ossia la divisione del nucleo, alla quale segue la comparsa del primo solco sagittale il quale separa due blastomeri incompleti di destra e di sinistra. Nella seconda divisione appare anche il secondo solco sagittale perpendicolare al primo, che separa due blastomeri anteriori e posteriori. Nelle terza divisione i due blastomeri anteriori sono divisi in maniera eguale a discapito di quelli posteriori che, nella quarta divisione, si separano in due cellule ciascuno. I blastomeri che rimangono aperti sul deutoplasma prendono il nome di blastoconi o cellule marginali. Le cellule marginali, durante le successive divisioni, formano il sincizio perilecitico che ha il compito di elaborare il deutoplasma.
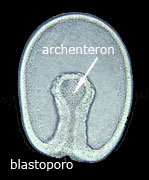
Le cellule della blastula sono chiamate blastomeri, mentre la cavità interna prende il nome di blastocele. Nella gastrulazione la blastula, mediante invaginazione, si trasforma in gastrula.
Il celome è una cavità del corpo rivestita da mesoderma e detta secondaria in contrapposizione al blastocele. Tipicamente il celoma si forma per schizocelia nei Protostomi e per enterocelia nei Deuterostomi. Nella condizione primitiva il celoma è costituito da una sequenza di sacche, una coppia per metamero, disposte lateralmente all'intestino ed estese fino alla parete del corpo. Dorsalmente e ventralmente le pareti dei comparti celomatici delle singole coppie formano i mesenteri dorsale e ventrale; la parete mediale, che avvolge l'intestino, e quella laterale, addossata alla parete del corpo, sono dette rispettivamente splancnopleura e somatopleura. Le varie sacche possono fondersi e formare compartimenti celomatici più grandi. La cavità celomatica è ripiena di liquido in cui navigano elementi cellulari (celomociti) o formazioni pluricellulari e ha funzioni di trasporto e distribuzione. Tranne che nei Platelminti e nei Nemertini, il celoma si forma probabilmente in tutti gli animali che presentano mesoderma, anche se pụ andare incontro a riduzioni di vario grado, fino alla scomparsa quasi totale. Per esempio, nei Policheti e negli Oligocheti il celoma è ben sviluppato e conformato in modo tipico, mentre in alcuni Irudinei pụ essere ridotto a una serie di lacune celomatiche. Gli Artropodi, parimenti, hanno celomi ridotto alle sole cavità delle gonadi (gonocele), delle ghiandole antennali e coxali (nefrocele), e la cavità del corpo (emocele) si identifica con il blastocele. Nell'embrione dei Cordati il celoma si forma nello spessore delle piastre laterali del mesoderma per delaminazione di questo in due foglietti. Inizialmente il celoma è costituito da due cavità simmetriche bilaterali, internamente rivestite da un mesepitelio pavimentoso (celotele), disposte una per parte ai lati dell'intestino che ne risulta circondato. Dorsalmente e ventralmente all'intestino le cavità celomatiche di destra e di sinistra vengono a contatto formando i mesenteri dorsale e ventrale. Nel rivestimento delle cavità celomatiche si distinguono un foglietto viscerale che aderisce all'intestino e forma i mesenteri, detto splancnopleura, e uno parietale che tappezza le pareti interne del corpo, detto somatopleura. Splancnopleura e somatopleura continuano anche al di fuori del corpo dell'embrione fornendo di un rivestimento mesoteliale anche gli eventuali annessi embrionali (corion, sacco del tuorlo, amnios e allantoide) e delimitando una concamerazione della cavità celomatica detta celoma extraembrionale. Nelle susseguenti fasi di sviluppo il celoma extraembrionale si separa dalla restante parte interna all'embrione. Quest'ultima, in seguito alla scomparsa del mesentere ventrale, da doppia diventa unica e prende il nome di cavità pleuroperitoneale. Nella parte di questa più vicina alla testa viene poi a formarsi il sacco pericardico e inoltre, nei Mammiferi, con la formazione del diaframma che suddivide trasversalmente il celoma, si separa la cavità pleurica da quella peritoneale. Per quanto riguarda il celoma gli animali possono distiguersi in: ACELOMATI, PSEUDOCELOMATI, CELOMATI (Tentacolati, Schizocelomati, Enterocelomati). Gli acelomati non possiedono un celoma (sono diblastici); gli pseudocelomati possiedono un falso celoma rappresentato dalla cavità virtuale tra ectoderma, entoderma e cellule mesodermiche sparse, non organizzate in un vero e proprio foglietto mesodermico; i tentacolati (Foronoidei, Briozoi, Brachiopodi) sono caratterizzati da un celoma di origine particolare, laddove cellule mesodermiche libere migrano con movimenti ameboidi disponendosi, quindi, in uno strato peritoneale continuo; negli schizocelomati (anellidi, molluschi, artropodi) il celoma si forma a partire da due cellule di derivazione endodermica, ciascuna ai lati del futuro intestino; infine, negli enterocelomati (echinodermi, cordati) il celoma si origina da tasche laterali che si estroflettono dall' endoderma.

Alla fine della gastrulazione l'embrione già possiede un intestino primitivo e strati di cellule che porteranno successivamente alla formazione di tutti i tessuti ed organi del corpo.
L'organogenesi, infine, è il processo con cui si formano i definitivi tessuti ed organi e si determina l'accrescimento corporeo dell'embrione.
In particolare, dall’ectoderma si formano l’epidermide, il sistema nervoso, gli organi di senso;
dall’endoderma si formano il rivestimento interno dell’apparato digerente e le ghiandole annesse (fegato e pancreas);
dal mesoderma si formano il rivestimento della principale cavità del corpo (il celoma), la muscolatura, l’apparato circolatorio, il sangue, l’apparato respiratorio e, nei vertebrati, lo scheletro e l’apparato urogenitale.
Nei rettili, uccelli e mammiferi lo sviluppo embrionale è caratterizzato anche da aree extra-embrionali, dette "annessi embrionali". Si tratta dell ' amnios, corion, sacco vitellino, allantoide e placenta, quest' ultima presente nei soli mammiferi placentati. 
Gli annessi embrionali sono strutture necessarie per l' adattamento di questi vertebrati alla vita in ambiente terrestre, in particolare servono per la protezione, per l' alimentazione come pure forniscono l' ambiente (acquatico) indispensabile per lo sviluppo (amnios). L' amnios è costituito da un sacco che si sviluppa a carico di una parte dei tessuti formati dall’uovo fecondato (sacco amniotico); contiene, in quantità variabile a seconda delle diverse specie animali e dei vari stadi di sviluppo, un liquido sieroso (liquido amniotico) nel quale si trova immerso l’embrione. Il liquido amniotico protegge l’embrione da eventuali urti e variazioni di pressione, ne facilita i movimenti e ne impedisce l’essiccamento. Inoltre, l' amnios ed il liquido amniotico in esso contenuto rappresentano una sorta di retaggio filogenetico del fatto che la vita si sarebbe originata in ambiente acquatico (marino); l' ontogenesi, pertanto, ripercorrerebbe in tal modo le prime tappe dell' evoluzione (Legge biogenetica fondamentale o della ricapitolazione). Per la presenza di amnios i rettili, gli uccelli ed i mammiferi vengono detti anche amnioti, in contrapposizione a tutti gli altri organismi che non necessitano e non hanno amnios e che, pertanto, vengono detti anamni. Il corion è costituito da una membrana che avvolge l’ embrione e delimita con la propria parete la cavità extraembrionaria. Ha un’importante funzione nutritiva mettendo in rapporto l’embrione con la madre. Tale rapporto si stabilisce tramite proliferazioni vascolarizzate ( villi coriali), che si formano sulla superficie del c̣rion e si insinuano nello spessore della mucosa uterina con la quale contraggono rapporti più o meno intimi dando origine a diversi tipi di placenta. I villi si formano su tutta la superficie del c̣rion e in seguito possono permanere tutti o regredire in parte. Il sacco vitellino è una piccola "vescica" che fornisce il nutrimento necessario all' embrione. Esso trae origine dalla primitiva cavità lecitocelica da cui si forma pure l'intestino primitivo. Nei Mammiferi euteri ha dimensioni ridotte e si presenta come un sacco disposto attorno ad una cavità che contiene liquido , mentre nello spessore della paret del sacco si trovano vasi appartenenti al circolo vitellino. Tramite i vasi del circolo vitellino arrivano all'embrione i materiali nutritizi e l' ossigeno prelevati dal corion. Il sacco vitellino nella fase iniziale del suo sviluppo, stabilisce dei rapporti col corion, formando la c.d. onfalo-placenta; esso è comunque destinato a regredire a mano a mano che si sviluppa l'allantoide. Inoltre, il sacco vitellino, all'inizio, si presenta variamente sviluppato nelle diverse specie di Mammiferi euteri: nei Ruminanti non stabilisce mai dei rapporti col corion, regredisce progressivamente ed alla nascita non rimane più traccia; negli Equidi si forma un'onfalo-placenta vera, ma transitoria per la sua durata (è presente soltanto fino al quarto mese di gravidanza); nei Carnivori i rapporti onfalo-placentari persistono sino alla nascita.
Negli animali in cui il nuovo individuo nasce dal corpo della madre, si parla di viviparità; quando invece esso si schiude da un uovo, deposto nell’acqua o sul terreno, si parla di oviparità. In alcune specie animali si verifica anche il fenomeno della ovoviviparità; il nuovo individuo, cioè, fuoriesce da un uovo, che è rimasto nelle vie genitali femminili e, pertanto, prosegue il suo sviluppo all’interno del corpo materno. Cị avviene, ad esempio, nella lucertola Lacerta vivipara e nella vipera Vipera berus. In alcuni animali a sviluppo indiretto, dal cui uovo fuoriesce una larva, tale processo pụ svolgersi anche all’interno della madre; questa, dunque, “partorisce” delle larve. In tal caso, si parla di larviparità, fenomeno noto in alcune salamandre, nel coleottero Chrysomela e nell’insetto efemerottero Cloëon.
Le cellule staminali sono cellule in grado di mantenere capacità proliferative durante tutta la vita dell'individuo; esse si dividono asimmetricamente, con una delle due cellule figlie che rimane di tipo staminale e l'altra che inizia il processo di differenziamento. Le cellule staminali possono distinguersi in totipotenti, capaci cioè di trasformarsi in qualsiasi tipo di tessuto, pluripotenti, che si trosformano solo in alcuni tipi di tessuti, e unipotenti, che possono dar luogo soltanto ad un tipo cellulare. Nei mammiferi, le cellule staminali totipotenti sono presenti nella massa di cellule del nodo embrionale della blastocisti nelle fasi di preimpianto, nell'embrione e nel feto durante lo sviluppo e si ritrovano persino nell'individuo adulto, anche se solo in alcuni precisi distretti dell'organismo (midollo spinale, epitelio seminifero della gonade maschile, retina, cervello). Se le cellule staminali di ciascuno di questi distretti vengono isolate e opportunamente coltivate, è possibile aumentarne il numero e differenziarle nel tipo cellulare specifico del distretto tissutale da cui derivano, o anche transdifferenziarle ottenendo coś, per esempio, cellule del sangue a partire da cellule staminali del tessuto nervoso. Una via alternativa per ottenere cellule staminali è il loro isolamento dal cordone ombelicale. Va comunque ricordato che le limitazioni numerica e fisiologica si presentano anche in questo caso, sebbene in minor misura. Negli ultimi tempi le cellule staminali, in particolare la loro creazione e il loro impiego in medicina, sono diventate il tema scientifico più attuale: le ricerche e le scoperte si susseguono a ritmo sfrenato, occupando non solo le pagine delle riviste scientifiche specializzate ma anche quelle dei quotidiani o dei settimanali più popolari. E sollevando più di una polemica: da una parte sulla capacità di queste cellule di curare malattie finora considerate inguaribili, dall'altra sulla moralità delle tecniche utilizzate per ottenerle.
All Rights Reserved. Text and images on this website may not be redistributed or put
   |