
"Anchialine (from Greek meaning "near the sea") refers to coastal caves formed in limestone or volcanic rock that are flooded with seawater. They include the longest submerged caves on Earth. These caves are inhabited by a diverse array of previously unknown species from a number of new higher taxa. While some are primitive "living fossils", others are closely related to deep sea species. Most lack eyes and pigment, owing to their existence in the perpetual darkness of underwater caves. While some closely related species are found in caves on opposite sides of the Atlantic Ocean, others, previously known exclusively from caves in the Atlantic Ocean, recently have been discovered in Western Australia. These highly irregular distributions suggest an origin many millions of years ago when the Earth's landmasses were interconnected. Since such anchialine cave animals are frequently limited to a single cave or cave system, pollution or destruction of these caves can result in the extinction of entire species". (from: www.tamug.edu/cavebiology) I sistemi idrici di molte grotte marine a di altri ambienti sotterranei litorali costituiscono particolari biotopi separati da quelli costieri circostanti da forti gradienti ambientali. In questi habitat, definiti «anchialini» (dal greco = vicino al sale, mare), le particolari condizioni edafiche, idrografiche ed idrologiche hanno favorito il diversificarsi di ricche biocenosi stigobionti e lo sviluppo di faune endemiche di rilevante interesse scientifico. La loro scoperta si deve a Riedl (1966) che li denominņ "Randhoehlen" o "marginal caves", limitandone, tuttavia, la definizione alle sole grotte marine. Molti anni pił tardi Holthuis (1973), in occasione del rinvenimento di alcuni interessanti decapodi in ambienti sotterranei costieri indopacifici, ripropone il problema di questi interessanti biotopi attribuendo loro per la prima volta il nome di "anchialine pools" e definendoli come corpi idrici sotterranei senza connessioni superficiali con il mare, caratterizzati da acque salate e salmastre fluttuanti con le maree ("pools with no surface connection to the sea, containing brackish water, which fluctuates with the tides"). Successivamente nuovi termini sono stati introdotti, tuttavia non sempre in modo appropriato, per definire questi ambienti: "inland marine caves", "sea water-flooded caves", "anchihaline cave waters", "anchihaline habitats", "metahaline anchihaline pools", "grottes de dissolution" (Iliffe 1991; Stock, 1994; Sket, 1996; Juberthié & Decu, 1994). Bisogna attendere sino al 1984 quando, in occasione del Congresso Internazionale di Biologia delle grotte marine, tenutosi in Bermuda, si stabili definitivamente che con il termine «ambienti anchialini» dovessero essere considerati quei corpi idrici a salinita variabile, poco esposti all' aria aperta, con pił o meno estese comunicazioni sotterranee con il mare e con evidenti influenze marine e continentali ("Anchaline habitats consist of bodies of haline waters, usually with a restricted exposure to open air, always with more or less extensive subterranean connections to the sea, and showing noticeable marine as well as terrestrial influences"). In definitiva gli habitat anchialini sarebbero rappresentati da quei sistemi idrici sotterranei (cavernicoli, freatici, di frattura carsica) situati in prossimitą di coste marine, ad esse strettamente collegati, le cui caratteristiche principali sarebbero: tipologia di habitat sotterraneo, basso input energetico, assenza di luce, acque mixoaline, ipossiche o addirittura anossiche, mancanza di connessioni superficiali con il mare, limitato accesso di fauna marina e presenza di caratteristici organismi stigobionti con scarso potenziale dispersivo. Inoltre, l'assenza di turbolenze in tali ambienti impedirebbe il rimescolamento delle acque marine e carsiche con diversa densita le quali si stratificherebbero, quindi, in livelli diversi, dando origine a gradienti verticali di temperatura, salinitą e di concentrazione dell'ossigeno disciolto. In molti casi č possibile individuare un netto livello di separazione tra acque dolci e salate (haloclino) influenzato dalla distanza dal mare, dalla piovositą e dalla geomorfologia del territorio. In alcuni casi, tuttavia, moto ondoso ed abbondante piovosita producono turbolenze che possono alterare temporaneamente la suddetta stratificazione, annullando il relativo haloclino.  Gradienti di salinitą e temperatura nella grotta Zinzulusa (Otranto, Lecce) Esiste una grande varietą di ambienti anchialini: depressioni idriche sotterranee quali, ad esempio, le pozze anchialine che si rinvengono in terreni lavici molto porosi nelle Hawaii; faglie tettoniche costiere in rocce calcaree (Pacifico centrale) o laviche (Galapagos) al di sotto del livello del mare; grotte calcaree parzialmente o totalmente sommerse (Caraibi, Bahamas, Bermuda, Mediterraneo); sistemi freatici costieri (pozzi) in substrati calcarei (Italia meridionale) o lavici (Antille, Australia) (Humphreys, 1993)
Particolari manifestazioni anchialine sono quelle rappresentate da sistemi idrici all'interno di piccole fessure o fratture in rocce vulcaniche ("lava tubes") o calcaree (Isole Canarie, Galapagos, Hawaii, Samoa) (Humphreys, 1993; Iliffe, 2000), i cosidetti "blue holes" delle Bahamas, consistenti in fratture sottomarine circolari sottoposte a forti correnti di marea che spingono ciclicamente le acque marine all'interno e all'esterno delle cavitą, e alcuni "cenotes" della penisola dello Yucatan (Messico). Questi ultimi ambienti, molto ben rappresentati lungo le coste caraibiche del Messico, sono costituiti da cavitą, spesso molto lontane dalla costa, contenenti acqua marine e dolce, stratificate secondo forti gradienti di salinitą ("haloclines"). La maggior parte degli ambienti anchialini č caratterizzata da una elevata diversita biologica. I taxa maggiormente rappresentati sono i crostacei (remipedia, termosbenacei, anfipodi, ostracodi, copepodi, isopodi cirolanidi, mictacei, bochusacea, misidacei, tanaidacei, decapodi, fillocaridi), meno frequenti risultano batteri, protozoi, idrozoi, ciliofori, poriferi, anellidi (policheti ed oligocheti), molluschi, chetognati e pesci. Dei suddetti taxa alcuni gruppi di crostacei (Atlantasellidae, Remipedia, Platycopioida, Speleoithonidae, Mictacea) risultano esclusivi degli ambienti anchialini. In particolare i copepodi ciclopoidi, calanoidi, misofrioidi ed arpacticoidi, sembrano essere i piu comuni e numerosi colonizzatori dei sistemi anchialini dell' area mediterranea a dell' Atlantico orientale. A tutt'oggi oltre 250 nuove specie, 50 nuovi generi, almeno 10 nuove famiglie, due nuovi ordini ed una nuova classe di crostacei (Remipedia) sono stati descritti per gli ambienti anchialini, soprattutto di grotte insulari. In particolare la recente scoperta dei Misofrioida in grotte anchialine delle Bermuda ha messo in evidenza la possibilitą per i rappresentanti di questo gruppo di crostacei, un tempo ritenuti esclusivi di acque molto profonde, di colonizzare anche sistemi idrici meno profondi, costieri (Boxshall, 1989). La maggior parte dei taxa anchialini pił specializzati esibiscono caratteristiche stigomorfie, quali la riduzione degli organi visivi, la depigmentazione, l'aumento della sensibilitą tattile e chimica ed il rallentamento dello sviluppo, con produzione di pochissime uova. Una totale depigmentazione e perdita degli occhi si riscontrano in numerosi taxa anchialini, per lo pił in forme molto primitive che sono state relegate per lungo tempo nell'ambiente sotterraneo (anellidi policheti, remipedia, ostracodi, copepodi misophrioidi, isopodi cirolanidi, termosbenacei, anfipodi ed alcuni pesci). Per le loro caratteristiche molto ancestrali, una gran parte dei crostacei anchialini (remipedia, copepodi dei generi Antrisocopia e Erebonectes, platycopioida, misophrioida) vengono considerati veri e propri "fossili viventi" (Iliffe, 2000). Poichč la maggior parte delle specie viventi in ambienti anchialini sono frequentemente limitate ad una sola grotta o cavitą, l' inquinamento o la distruzione di tali habitat potrebbe determinarne l' estinzione!
ANCHIALINE ATTUALMENTE CONOSCIUTE
Gli stigobionti anchialini sono quasi esclusivamente di origine marina e consistono per la maggior parte di crostacei; inoltre numerosi stigobionti dulciacquicoli (turbellari, nematodi, anellidi policheti, copepodi, misidacei, anfipodi, isopodi, termosbenacei e pesci) sono relitti marini che hanno colonizzato le acque dolci sotterranee tramite habitat anchialini. Nonostante esse siano generalmente confinate in ambienti cavernicoli, la gran parte delle specie anchialine presentano ampie distribuzioni geografiche, in molti casi (remipedia, copodi misophrioidi, termosbenacei, misidacei, ostracodi thaumatocypridi) fortemente disgiunte, che pongono il problema della relativa origine. Una grande varietą di generi contiene specie a distribuzione anfi-atlantica, cioč localizzate in grotte sulle opposte coste dell'Oceano Atlantico, e di origine Tetidiana (remipedia, anfipodi, misidacei, decapodi), altri generi precedentemente conosciuti solo per grotte dell'area Caraibica, sono stati recentemente rinvenuti nelle Galapagos. Pił complesse relazioni biogeografiche esistono per altri taxa anchialini, tra cui i decapodi del genere Procaris conosciuti per le Hawai, le Bermuda, l'isola di Ascension e l'area Caraibica. Sorprendenti sono, infine, i casi dei misophrioida del genere Expansophria e dei misidacei del genere Palaumysis, con specie anchialine presenti a Palau, Galapagos, Bahamas e Canarie (Iliffe, 2000; Pesce & Iliffe, 2000). Infine taxa anchialini (remipedia del genere Lasionectes, termosbenacei del genere Halosbaena, ostracodi del genere Danielopolina ed anfipodi del genere Liagoceradocus) , ritenuti limitati all'area atlantica e Caraibica, sono stati recentemente scoperti in grotte e sistemi freatici dell'Australia nord-occidentale (Cape Range Peninsula) (Humphreys, 1993). L'ampia distribuzione di tutti i suddetti taxa, e di molti altri ancora, suggerisce per la maggior parte di essi una notevole antichitą, correlabile con meccanismi di dispersione simili alla colonizzazione degli habitat attuali. Un rilevante numero di specie anchialine, appartenenti a gruppi diversi quali Decapodi, Anellidi Policheti, Mictacei, Ostracodi, anfipodi e misofrioidi, presentano strette affinitą con forme marine batiali. Numerose e tuttora molto controverse sono le teorie e le ipotesi suggerite per interpretare tutte le suddette distribuzioni e per spiegare la origine delle faune anchialine. Una origine da acque profonde ("deep-sea origin") č stata invocata per numerosi gruppi di crostacei (Galatheidi, Mictacei, anfipodi Pardalascidi, ostracodi del genere Danielopolina, misiphrioidi) che si sarebbero evoluti a partire da antenati marini batiali tramite dispersione attraverso un "continuum" di sistemi crevicolari ("crevicular habitats") che avrebbero, appunto, costituito un link tra le acque pił profonde ed i sistemi anchialini costieri (Iliffe et al., 1983; Korniker & Iliffe, 1985; Boxshall, 1989; Boxshall & Jaume, 2000). Una interessante alternativa al suddetto modello di colonizzazione č l'ipotesi di una origine Tetidiana ("Tethyan hypothesis") applicabile a taxa (copepodi Stephidi e Pseudocyclopinidi, Speleophrioidi, termosbenacei del genere Halosbaena, remipedia dei generi Lasionectes e Speleonectes, misidacei dei generi Spelaeomysis e Stygiomysis) con evidente distribuzione tetidiana, i quali non hanno mai raggiunto gli habitat pił dolci, interni dei sistemi anchialini. secondo questa ipotesi i modelli distribuzionali dei suddetti taxa porterebbero alla conclusione di considerarli relitti di una fauna calda tetidiana, un tempo molto pił ampiamente distribuita. Le diverse modalitą con cui queste forme avrebbero colonizzato gli ambienti anchialini vengono sintetizzate nei modelli "Regression Model" (Stock, 1977, 1980, 1990: Boutin & Coineau, 1990), "active Migration Model" e "Passive Migration Model (Rouch & Danielopol, 1987). Il modello pił comune (Regression Model) si basa sulla osservazione che i patterns di distribuzione di molti taxa anchialini sono perfettamente correlabili con le linee di costa dei mari mesozoici e terziiari. Per interpretare tali distribuzioni il "Regression Model" suggerisce che specie marine litorali avrebbero primariamente colonizzato habitat crevicolari ed interstiziali, quindi , a seguito delle regressioni marine, sarebbero rimasti "intrappolati" e adattati in ambienti mixoalini, in acque limniche sotterranee. Tali migrazioni potrebbero essere avvenute sia tramiteprocessi attivi (Active Migration Model) che passivi (Passive Migration Model ) Stock, 1986; Boxshall & Jaume, 2000). . Un particolare scenario deve essere proposto per quanto riguarda l'origine degli stigobionti anchialini mediterranei, in gran parte riconducibile alla cosidetta «crisi di salinita messiniana» realizzatasi circa 5.5 milioni di anni fa allorquando, in accordo con l'ipotesi di Hsu (1978), il Mediterraneo si sarebbe quasi completamente prosciugato con conseguente parziale scomparsa della relativa fauna. Recenti studi di Sket (1996) e Boxshall & Evstigneeva (1994) hanno, infatti, dimostrato che un gran numero di stigobionti mediterranei attuali (copepodi Mysophriidae, Pseudocyclopiidae, Cyclopidae; misidacei; anfipodi; remipedia; decapodi; spugne), con geonemie disgiunte, presenti in habitat ipogei con salinitą prossima a quella marina (>18 x mille) sarebbero derivati appunto da antenati marini sopravvisuti alla suddetta crisi grazie alla loro elevata eurialinita. In conclusione, la combinazione di un modello di distribuzione tetidiano con la loro localizzazione in ambienti ad elevata salinita rappresenta la dimostrazione evidente di come molti stigobionti anchialini sarebbero sopravvissuti in situ nel corso del Terziario, un periodo che vide, appunto, il Mediterraneo scenario di drammatici eventi ambientali che ne comportarono il quasi completo prosciugamento e la relativa scoparsa di gran parte dei suoi biotopi. In ogni caso, si deve mettere in evidenza la rilevante importanza degli ambienti anchialini quali "rifugi" di antiche linee evolutive, spesso rappresentate da singole specie limitate ad una sola cavitą anchialina, come pure riconoscere che l'estrema "fragilitą" ecologica di questi habitat richiederebbe pił adeguate misure di protezione e tutela al fine di evitare la possibile estinzione di specie o di interi gruppi animali di eccezionale valore scientifico. Ambienti con caratteristiche anchialine sono attualmente noti per numerose localita in quasi tutto il mondo: Hawaii, Bermuda, Bahamas, Galapagos, Messico, Antille, Canarie, Pacifico centrale, isole Fiji e Ryukyus, Sinai (Mar Rosso), Oceania, Australia, coste adriatiche della Croazia e Dalmazia, Grecia, Baleari, Italia. 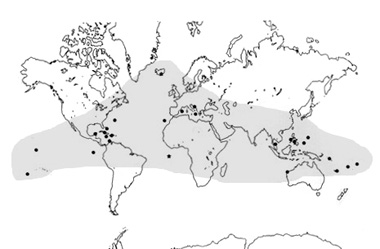 Mappa mondiale delle principali aree anchialine (da Iliffe, 2000, mod.) In Italia numerose, anche se poco studiate, sono le manifestazioni anchialine distribuite lungo le coste della Toscana, Gargano, Campania, Puglia, Sicilia e Sardegna. Da un punto di vista generale, i sistemi anchialini italiani risultano densamente popolati, soprattutto da crostacei (Pseudponiphargus, Microcharon, Monodella, Spelaomysis, Typhlocaris, Hadzia, Stygiomysis, Orniphargus, Salentinella, Diacyclops, Halicyclops, Neocyclops, Schizopera), spugne, idracari ed anellidi oligocheti. Attualmente, molto ben conosciuti risultano i popolamenti anchialini della Puglia (Gargano, Penisola Salentina) e della Sicilia, grazie ad una serie di intensive ricerche condotte dal Dipartimento di Scienze Ambientali dell' Universitą di L'Aquila, con la collaborazione di gruppi speleologici delle due regioni.
In Puglia gli ambienti anchialini risultano ben rappresentati sia all'interno delle grotte costiere della penisola Salentina che nei sistemi freatici litorali, situati a pochi metri di distanza dal mare, lungo i litorali adriatico e ionico.
Da un punto di vista biologico, la grotta della Zinzulusa mostra il pił elevato numero di specie, oltre 60 , la maggior parte delle quali stigobionti ed endemiche per la grotta o per il distretto salentino. Tra queste alcune appartengono a generi (Spelaeomysis, Stygiomysis, Hadzia) presenti anche nell'area Caraibica, altre a generi (Typhlocaris, Monodella, Salentinella, Lohmannella, Soldanellonix) con geonemie pił limitate, alcune, infine, risultano endemiche per il territorio Salentino. L'elemento, comunque, pił sorprendente, recentemente scoperto all'interno della grotta, č la spugna troglobia "Higginsia ciccaresei" . Si tratta di uno dei rarissimi reperti di spugne in ambienti sotterranei, anchialini, con elevate stigomorfie; a tutt'oggi, infatti, gli unici riferimenti a spugne ipogee, peraltro stigofile, erano quelli relativi a rinvenimenti in grotte marine del Mediterraneo (Vacelet & Boury-Esnault, 1995) delle Bahamas (Van Soest & Sass, 1981; Iliffe, 2000) . Per il suo eccezionale interesse biologico la grotta Zinzulusa č stata recentemente inclusa nella "Top ten list of World Endangered Karst Ecosystems" dal "Karst Water Institute" Charles Town WV, U.S.A., rappresentando la pił importante manifestazione carsica anchialina dell'intero territorio italiano. Le grotte «Buco dei Diavoli» a «L'Abisso» sembrano essere caratterizzate da un minor numero di specie e da una bassa biodiversitą, comprendendo 11 e 12 taxa rispettivamente, la maggior parte dei quali presenti anche nella grotta Zinzulusa. Gli ambienti freatici (pozzi) lungo le coste adriatica e ionica esibiscono per la maggior parte caratteristiche di ambienti anchialini aperti, pił intimamente legati al mare. Si tratta, infatti, di pozzi molto prossimi alla costa, spesso in piena luce e con notevole apporto trofico, in cui risultano presenti componenti propriamente marine, di recente invasione, ed elementi pił specializzati penetrati nei sistemi freatici pugliesi in epoca pił antica (Pesce et al., 1978).
Tra questi ultimi si possono citare il misidaceo Spelaeomysis bottazzii, ampiamente distribuito, anche se con popolazioni ben caratterizzate, in tutto il territorio pugliese, il termosbenaceo Monodella stygicola, il decapode Typhlocaris salentina ed alcuni copepodi, sia ciclopoidi che arpacticoidi, anche questi ampiamente distribuiti nei diversi sistemi anchialini di tutta la Puglia.
Da un punto di vista biogeografico, la fauna pił propriamente stigobia dei pozzi risulta anch'essa sostanzialmente diversa e molto meno diversificata da quella presente nei sistemi anchialini di grotta. Essa presenta, infatti, accanto ad alcuni elementi endemici propri, quale l'isopode Microcharon arganoi, altre specie, ad ampia geonemia, presenti solo in questa parte della Puglia, tra cui gli anfipodi Salentinella angelieri a Bogidiella sp. ed il copepode ciclopoide Diacyclops clandestinus. D'altro canto in questa parte del territorio pugliese sembrano mancare, almeno a quanto ci č attualmente noto, relitti molto antichi, come pure alcuni dei pił caratteristici stigobionti presenti nei sistemi cavernicoli a freatici delta penisola Salentina. L'invasione dei diversi sistemi anchialini delta regione pugliese deve essersi realizzata tramite diverse «ondate di colonizzazione», alcune molto antiche di cui sarebbero testimonianza i numerosi relitti tetidiani (Higginsia ciccaresei, Monodella stygicola, Stygiomysis hydruntina, Typhlocaris salentina) sopravvissuti in habitat criptici ed isolati a da lungo tempo infeodati nei sistemi sotterranei dolci continentali. Infatti l'ipotesi pił «parsimoniosa», esplicativa delta distribuzione di generi quali Higginsia, Monodella, Typhlocaris e Stygiomysis, sembra essere quella di considerarli appunto relitti di una fauna litorale antica (tardo Mesozoico), un tempo ampiamente distribuita in acque tropicali a subtropicali come dimostrato dalla perfetta coincidenza tra le loro attuali geonemie a le antiche linee di costa mesozoiche del Mediterraneo. Alcune specie (Hadzia minuta, Microcharon arganoi) avrebbero migrato negli stessi sistemi in tempi pił recenti, in occasione della ipotizzata «crisi di salinitą» the caratterizzņ il Mediterraneo durante il Miocene; altre, tra cui il misidaceo Spelaeomysis bottazzii a gli anfipodi Salentinella angelieri e Salentinella gracillima, avrebbero seguito gli spostamenti delle linee di costa del Mediterraneo durante il Pliocene. Alcuni copepodi arpacticoidi, quali Esola spelaea, Psyllocamptus monachus, Schizopera clandestina a Schizopera cicolanii e l'anflpode Pseudoniphargus adriaticus, rappresentano un esempio di forme marine immigrate, in tempi relativamente recenti, nelle acque sotterranee continentali dopo un lungo periodo di preadattamento nei sistemi carsici litorali, molto probabilmente in accordo con il «two-step model of colonization» suggerito da Boutin & Coineau (1990). Per quanto rigurda, infine, la Sicilia le uniche informazioni attualmente disponibili si riferiscono ai sistemi anchialini dell'area carsica di Porto Palo. In questa zona numerosi risultano i pozzi costieri all'interno dei quali č presente una stigofauna dalle evidenti caratteristiche anchialine, comprendente numerosi copepodi, ciclopidi (Halicyclops troglodytes, Eucyclops ibleicus, Eucyclops longispinoisus, Diacyclops clandestinus, Diacyclops crassicaudis trinacriae) ed arpacticoidi (Nitocrella stammeri Attheyella paranaphtalica, Elaphoidella elaphoides, Elaphoidella rossellae, Parastenocaris trinacriae), anfipodi (Niphargus longicaudatus), termosbenacei, acari alacaridi (Soldanellonyx monardi, Porohalacarus alpinus, Lobohalacarus weberi), isopodi asellidi (Proasellus coxalis), ostracodi, gasteropodi ed anellidi oligocheti. La fisionomia ecologica della stigofauna anchialina di questa parte della Sicilia risulta molto interessante. Accanto ad una bassa percentuale di elementi stigosseni, ad ampia adattabilitą, figurano infatti numerosi rappresentanti pił specializzati, alcuni propriamente stigobionti e di antico insediamento nelle biocenosi anchialine della regione (Pesce & Galassi, 1987), altri, "talassoidi", di pił recente invasione nei biotopi sotterranei che avrebbero colonizzato in concomitanza dei cicli geocratici del Mediterraneo durante il Mio-Pliocene, secondo il "Regression Model Evolution" suggerito da Stock (1977, 1980). Boutin C. & N. Coineau. 1990. "Regression Model", "Modele biphase" d'evolution et origine des micro-organismes stygobies interstitiels continentaux. Rev. Micropaleontol., 33: 303-322. Boxshall G.A. 1989. Colonization of inland marine caves by misophrioid copepods. J. Zool., 219: 521-526. Boxshall G.A. & T.D. Evstigneeva. 1994. The evolution of species flocks of copepods in Lake Baikal a preliminary analysis. arch. Hydr. Balh. Limn., 44: 235-245. Boxshall G.A. & D. Jaume. 2000. Discoveries of cave Misophrioids (Crustacea: Copepoda) shed new light on the origin of anchialine faunas. Zool. Anz., 239: 1-19. Holthuis L.B. 1973. Caridean shrimps found in land-locked saltwater pools at lour Indo-West pacific localities (Sinai Peninsula, Funafuti atoll, Maui and Hawaii islands), with the description of a new genus and four new species . Zool. Verh., 128: 1-48. Hsu K.J. 1978.when the Black sea was drained. Sci. Amer., 238, 5: 53-63. Humphreys W.F. 1993. Stygofauna in semi-arid tropical Western Australia: a Tethyan connection? . Mem. Biospeol., 20: 111-116. Iliffe T. M. 1991. Anchialine fauna of the Galapagos Islands. in: Galapagos Marine Invertebrates. J. M.J. ed., pp.209-211. Iliffe T. M. 2000. Anchialine cave ecology. in: Ecosystems of the World 30. Subterranean Ecosystems. Elsevier ed. pp.59-76. Iliffe T. M. , C. W. Hart & R. B. Manning. 1983. Biogeography and the caves of Bermuda. Nature, 302: 141-142. Juberthič C. & V. Decu. 1994. Structure et diversite du domain souterrain: particularites des habitats et adaptations des especes. in: Encyclopaedia Biospeleologica, Moulis-Bucarest, pp. 5-22. Kornicker L.S. & T.M. Iliffe. 1985. Deeveyinae, a new subfamily of ostracoda (Halocyprididae) from a marine cave on the Turks and Caicos Islands. Proc. Biol. soc. Wash., 98: 476-493. Pansini A. & G. L. Pesce. 1998. Higginsia ciccaresei sp. nov. (Porifera: Demospongiae) from a marine cave on the Apulian coast (Mediterranean Sea). J. Mar. Biol. Ass. U.K., 78: 1083-1091. Pesce G.L. & T.M. Iliffe. 2000. New records of cave-dwelling mysids from the Bahamas and Mexico with description of Palaumysis bahamaensis n.sp. (Crustacea: Mysidacea). J. nat. Hist. (in press). Pesce G.l. , G. Fusacchia, G. Maggi & P. Tetč. 1978. Ricerche faunistiche in acque freatiche del salento. Thalassia Salentina, Lecce, 8: 1-51. Riedl R. 1966. Biologie der Meershoelen. P. Parey ed., pp.1-23. Rouch R. & D.L. Danielopol. 1987. L'origine de la faune aquatique souterraine, entre le paradigme du refuge et le modele de la colonisation active. Stygologia, 3: 345-372. Sket B. 1996. The ecology of anchialine caves. Trends in Ecology and Evolution, 11 (5): 221-224. Stock J. H. 1977. The taxonomy and zoogeography of the hadziid Amphipoda, with emphasis on the West Indian taxa. stud. Fauna Curacao, 177: 1-130. Stock J. H. 1980. Regresion model evolution as exemplified by the genus Pseudoniphargus (Amphipoda). Bijdr. Dierk., 50: 105-144. Stock J. H. 1986. Deep sea origin of cave faunas, an unlikely supposition. Stygologia, 2: 105-111. Stock J. H. 1990. Insular groundwater biotas in the (sub) tropical Atlantic: a biogeogaphic synthesis. atti Conv. Lincei (acad. Naz. Lincei, Roma), 85: 695-713. Stock J. H. 1994. Biogeographic synthesis of the insular groundwater faunas of the (sub) tropical Atlantic. Hydrobiologia, 287: 105-117. Vacelet J. & N. Boury-Esnault. 1995. Carnivorous sponges. Nature, 373 (6512): 333-335. Van Soest R.W.M. & D.B. Sass. 1981. Marine sponges from an island cave on San Salvador Island, Bahamas. Bijdr. Dierk., 51: 332-344.
All Rights Reserved. Text and images on this website may not be redistributed or put
   |
